| 陸上植物は動き回ることができないため、環境変化に対して、柔軟でかつ適切な応答をしています。当研究室では、陸上植物の成長や生存に関係する光合成系や呼吸系、気孔調節が環境変化にどのように応答し、どのような生態学的な意義があるかを解明するために、モデル植物のシロイヌナズナやフラベリア、薬用植物、野外の植物を使って研究しています。 |
|
| (1) CO2吸収とH2O損失のバランスを制御する仕組みの解明
|
| (2) 呼吸系と光合成系の相互作用 昼間の葉の呼吸系の役割
|
| (3) 葉の光合成系の季節変化をもたらすしくみの解明
|
| (4) その他
|
|
| 共同研究を積極的に進めています。研究内容について質問のある方や詳細が知りたい方、植物の呼吸系やガス交換について相談がある方は遠慮なく、野口(knoguchi@toyaku.ac.jp)もしくは溝上(ymzkm@toyaku.ac.jp)までご連絡ください。
|
|
|
|
| (1) CO2吸収とH2O損失のバランスを制御する仕組みの解明 |
| 陸上植物は光合成の基質であるCO2を葉の表面にある気孔から取り込みますが、同時に蒸散によって多くのH2Oを失います。環境の変化がCO2の取り込みとH2Oの損失のバランスを変えますが、成長を続けるために適切に制御されています。どのような制御機構があるのかを明らかにするために、気孔の制御、CO2取り込み能力、葉への水供給能力などを中心に研究を進めています。植物種として、シロイヌナズナの気孔応答性変異体やC3/C4型のさまざまな光合成を行うフラベリア属を扱っています。また、イチゴやその他の多くの植物種を用いて気孔以外の水の出口である水孔からの溢液(いつえき)現象にも注目して研究しています。
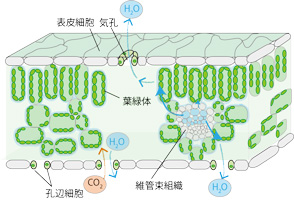  
左図: 葉の断面図 1分子のCO2を固定するために400分子のH2Oを失う
中図: サーモグラフカメラによりシロイヌナズナの葉温を測定した写真
気孔が開いている変異体は葉温が低い
右図:イチゴの溢液現象 鋸歯の先端に多く存在する水孔から排水される
|
|
|
| (2) 呼吸系と光合成系の相互作用 |
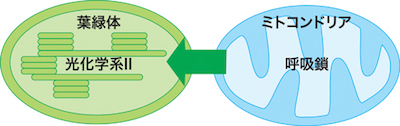
| 昼間に光を受けて光合成をしている葉では、葉緑体でおきる光合成とミトコンドリアでおきる呼吸との相互作用が重要だと考えられています。これまでに、シロイヌナズナの葉のミトコンドリア呼吸鎖を阻害した条件では、強光下で光合成の光化学系IIの活性や電子伝達系全体の活性が低下することを明らかにしました (Wataanbe et al. 2016, Jiang et al. 2019, Yamada et al. 2020)。現在、それらの低下要因を葉の分光学的測定や生化学的な実験を組み合わせて解析しています。今後はその生理学的機構の詳細や、植物種による相互作用の多様性やその意義を明らかにしていきたいと考えています。
|
|
|
|
| (3) 葉の光合成系の季節変化をもたらすしくみの解明 |
東京薬科大学キャンパス内に広がる落葉樹林の林床では、夏は薄暗く、冬から初春まで明るい季節変化があり、林床の草本植物種はそのような環境に適応しています。そのような植物種に絶滅危惧II類のタマノカンアオイがあります。1年の間に光強度や温度が大きく季節変動する環境で、タマノカンアオイは新しい葉を春に展開し、翌年の春に落葉する常緑草本種です。これまでに、タマノカンアオイの葉の光合成系は薄暗い夏に光合成系が抑制されていること、冬季の低温・強光というストレス環境下では、光合成速度が上がるともに光合成系を保護するような色素が誘導されることなどを明らかにしました(Wada et al. 2023)。
落葉樹林の林床にはタマノカンアオイ以外にも、林床が明るい春にだけ葉や花をつけるセツブンソウやキクザキイチゲのような春植物、春に葉を展開し、薄暗い夏までに葉を落とすウバユリのような夏植物、遅い秋に葉を展開し、冬を越して翌年の春に葉を落とすユキワリイチゲのような越冬性の葉をつける植物種など、葉をつける季節性(フェノロジー)が多様な草本植物種が見られます。異なるフェノロジーの植物種における光合成系の制御機構の違いを今後明らかにしていきたいと考えています。
 
常緑草本種タマノカンオアイ(左) 春植物セツブンソウ(右) タマノカンアオイでは地際に花弁のない花が見られる。
|
|
|
|
| (4) その他 |
| 薬用植物の研究(裸子植物で葉が退化しているマオウ科インテルメディアマオウの茎の光合成系の栄養条件への応答)、スイレンやアサザなどの水生植物の換気能力の解析、呼吸速度の温度依存性と成長との関連の解析、窒素栄養条件による光化学系の応答性の解析(Ozaki et al. 2022, 2023)、水陸両生植物種Hygrophila difformisの水中適応機構の解明などを進めています。
 
|